



Authors:
GAS Cruz1*, CA Ferreira-Neto1,2, AC Lira-Neto3 and RC Moura1*
1Institute of Biological Sciences, Pernambuco University, Recife, Brazil
2Genetics Department, Federal University of Pernambuco, Recife, Brazil
3Agronomic Institute of Pernambuco, Recife, Brazil
Received: 18 March, 2017; Accepted: 22 May, 2017; Published: 23 May, 2017
GAS Cruz / RC Moura, Institute of Biological Sciences, Pernambuco University, Recife, Brazil, E-mail:
Cruz GAS, Ferreira-Neto CA, Lira-Neto AC, Moura RC (2017) Isolation and Characterization of Microsatellite Markers in Canthon (Petelcanthon) Staigi (Coleoptera: Scarabaeidae) and Cross-Amplification in Related Species. Int J Agric Sc Food Technol 3(2): 032-035. DOI: 10.17352/2455-815X.000019
© 2017 GAS Cruz, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Dung beetles; Enriched library; Genetic diversity
The species Canthon (Peltecanthon) staigi is a Neotropical “dung beetle” from Brazilian Atlantic Rainforest, which plays a key ecological role reallocating organic material and in some cases being a seed disperser. Moreover, this species is considered a bio indicator once is sensible to environment changes like the habitat loss. Despite its’ ecological importance nothing is known regarding the population structure of this dung beetle species. Molecular markers are informative tools to evaluate the extent and distribution of genetic diversity of the C. (P.) staigi populations remaining. Here we report the isolation and characterization of the first microsatellite markers for C. (P.) staigi. Four polymorphic microsatellite loci were characterized with allele numbers ranging from four to five per locus. The observed and expected heterozygosis ranged from 0.466 to 0.516 and 0.485 to 0.623, respectively. All loci were observed on Hardy Weinberg equilibrium. Linkage disequilibrium was not detected in any loci. From a subset of 24 loci it was observed positive transferability of six loci on four different tribes of Scarabaeidae. The loci will be used for studying population genetic structure of C. (P.) staigi and the cross-species amplification success extends the utility of these markers to be applied also on related species.
Introduction
Canthon is a neotropical genus which belongs to Scarabaeidae (Scarabaeidae) subfamily and comprises about 170 species, commonly known as “dung beetles”. The species Canthon (Peltecanthon) staigi Pereira, 1953, is inserted in a well-defined telecoprid guilds of dung beetles and play a key ecological role in the reforestation, as part of process of reallocating organic material and seeds dispersal [1-3]. Besides that, C. (P.) staigi is a notable species due to its wide geographic distribution on different vegetation types of Atlantic forest between the states of Paraiba and Paraná in [4-8].
The C. (P.) staigi species is sensible to environmental changes like forest fragmentation, which disturb its population ecological dynamics on the level of guild structure, being considered a bio indicator [2,7,9-11]. Considering the high degree of Atlantic Forest fragmentation, the characterization of genetic diversity of C. (P.) staigi requires an understanding of the population genetic structure within and among the remaining forest fragments.
Co-dominant molecular markers such as microsatellites have been used as an informative tool to generate information on population genetics level of insects. However, there are no microsatellite markers developed for any species of Canthon genus. Here we report the isolation and characterization of four polymorphic microsatellite loci from C. (P.) staigi. Our data are informative for future studies of population genetic structure and also cross-amplification among related species of five tribes of Scarabaeidae subfamily.
Materials and Methods
The genomic DNA was isolated from two individuals of C. (P.) staigi species according to [12]. The isolated DNA was submitted to a digestion process by restriction enzyme AfaI (Promega, Madison, Wisconsin, USA) and the resulted fragments were linked to adapters (Rsa21 5’-CTCTTGCTTACGCGTGGACTA-3’ / Rsa25 5’-TAGTCCACGCGTAAGCAAGAGCACA-3’). The library was enriched for two monomers, (CT) 8 and (GT) 8, using biotinylated microsatellite probes. The capture of the target fragments was performed by the use of streptavidin-coated magnetic beads (Promega). The next step was to link the Microsatellite-enriched DNA fragments into a pGEM-T vector (Promega). After that the plasmids were introduced by electroporation into Escherichia coli XL-1 Blue strains. The transformed cells grew on petri dishes with LuriaeBertani (LB) agar medium containing X galactosidase (5-bromo-4-chloro-indolyl-B-D-galactoside) (20 mg/ml), IPTG (Isopropylthio-B-D-galactoside) (100 mM), ampicillin (50 mg/ml) and tetracycline (12,5 mg/ml).
A total of 96 recombinant colonies were obtained and sequenced using the Big Dye terminator version 3.1 Cycle Sequencing Kit (Applied Bio systems), T7 (5’-TAATACGACTCACTATAGGG-3’) and SP6 (5’-ATTTAGGTGACACTATAG-3’) sequencing primers and 3100 DNA Analyzer (Applied Bio systems). For clones containing SSR motifs, forward and reverse sequences were aligned to obtain the consensus sequence using Chromatogram Explorer 3.2 (Heraclea Bio Soft S.R.L., Romania). Microsatellite amplification-specific primers were designed from 24 loci of applicable flanking sequences with the software PRIMER3 [13]. For the polymorphic loci see table 1.
The polymerase chain reactions (PCR) were performed in a Bicycler 96-Well Thermal Cycler (Applied Bio systems) in a reaction volume of 10 µL containing 5 ng of DNA template, 10x PCR buffer, 0.50 mM MgCl2, 2.5 mM dNTP, 5 U/µL Taq polymerase (Invitrogen), 10 pmol forward primer and 10 pool reverse primer. A touchdown cycle program was used as follow: 94°C for 5 min, followed by 30 cycles of 94°C for 30 s, the annealing temperature for each primer for 45 s (The optimum annealing temperature of different primers was screened by gradient temperature PCR) and 72°C for 45 s, and a final extension of 72°C for 10 min. The amplicons were checked on 1,8% agarose gel stained with Gel Red (Biotium, Hayward, California, USA) and were genotyped in 7% polyacrylamide denatured gel stained with silver nitrate. The genotyping was performed using a 100bp DNA ladder (Invitrogen) as size standard marker. Sixty individuals from two populations collected on Atlantic forest fragments on the states of Paraiba and Pernambuco, Brazil, were analyzed regarding genetic diversity (Table 2).
-
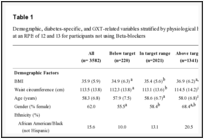
Table 2:
Location of sampled populations and parameters of genetic diversity in C. (P) staigi.
The 24 microsatellite loci were tested in seven species belonging to four tribes of Scarabaeidae subfamily. The PCR conditions and amplicons checking were performed as previously described. The bands amplified on the expected size were subsequently sequenced by Macrogen Inc (Seoul, Rep. of Korea), in order to check the presence of microsatellite motifs.
The genetic diversity per locus and population were evaluated on the basis of different indexes: allelic richness (R), number of alleles (A), observed (HO) and expected (HE) heterozygosis and inbreeding coefficient (FIS) on MSA program [14]. It was also calculated the polymorphic information content (PIC) [15] (Table 1). The Hardy-Weinberg equilibrium (HWE) and linkage disequilibrium (LD) were tested for all loci using Arlequin 3.5 [16]. Regarding the results robustness genotyping errors were calculated in relation to null alleles, stuttering, and allele dropout using Micro-Checker 2.2.3 [17].
Results and Discussion
The microsatellite enrichment library of 96 recombinant colonies resulted on 45 microsatellite repeat motifs, of which the most frequently encountered repeat motifs were (A)n, (AATT)n, (CTTTC)n, (TTGTG)n and (TGTGCA)n. The penta-nucleotides (71.11%) and hexa-nucleotides (17.77%) were frequently observed, followed by mono-nucleotides (6.66%) and tetra-nucleotides (4.44%). The di-nucleotides and tri-nucleotides were not found. The hexa-nucleotides (TTAAAA)n and (TGTGCA)n were found on the repetitive portion of the C.(P.) staigi genome related with a transposable element Tiger of the family Tc1/mariner, which is the most common on insect genomes [18].
From the 45 microsatellite 24 presented applicable flanking regions for primer design. From these four were polymorphic while 20 were monomorphic. The polymorphic loci generate a total of 19 alleles, ranging from four to five per locus. For monomorphic loci see table S1. The HO ranged from 0.466 to 0.516 while HE ranged from 0.485 to 0.623, per locus. The PIC index ranged from 0.312 to 0.368 per locus (Table 1). All loci were in HWE equilibrium and no linkage disequilibrium was detected. Furthermore, no evidence of scoring error was found.
-
Table S1:
List of loci which were monomorphic in Canthon (P) staigi (Scarabaeidae).
Regarding the two populations, GUA presented higher values of allelic richness, HE and HO than the PES population (Table 1). Interestingly, the observed FIS value in PES was lower than GUA, which means that population GUA could be suffering greater inbreeding depression, likely caused by geographic isolation on the range margin of the C. (P.) staigi spatial distribution.
The cross-species amplification tests revealed eight primer loci (Cstms02; Cstms06; Cstms10; Cstms12; Cstms14; Cstms15; Cstms19; Cstms20) which amplified satisfactorily among all species from four tribes. However, just six loci presented microsatellite motifs, indicating that are possibly useful for population genetics analysis of other beetle species (Table 3).
-
Table 3:
Transferability of eight microsatellite markers developed for C. (P) staigi accross five different tribes of Scarabaeinae (Scarabaeidae).
The four polymorphic loci isolated for C. (P.) staigi will be used to study the genetic diversity and population genetic structure of C. (P.) staigi providing knowledge which will be important for conservation strategies of this species.
Acknowledgments
We are grateful to MSc Cristiane Maria Quei- roz da Costa and Prof. Dr. Fernando Silva for the collection and iden- ti cation of specimens. This study was supported by a Grant from the Fundação de Amparo à Ciência e Tecnologia do Estado de Pernam- buco (FACEPE-IBPG-0831-2.02/11) through a Doctorate Scholar- ship to C.A.F.N., a Post-doc Researcher Fellowship to G.A.S.C. from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (PNPD-20130558) and FACEPE-BCT-0063-2.02/14, and by funding from the CNPq (Conselho Nacional de Desenvolvimento Cientí co e Tecnológico) to R.C.M. (CNPQ- 305298/2014-3).
- Halffter G, Favila ME, Halffter V (1992) "A comparative study of the structure of the scarab gild in Mexican tropical rain forests and derived ecosystems." Folia Entom Mex 84:131-156.
- Halffter G, Favila ME (1993) The Scarabaeidae (Insecta: Coleoptera) an animal group for analyzing, inventorying and monitoring biodiversity in tropical rainforest and modified landscapes. Biol Int 27: 15–21. Link: https://goo.gl/vU53yG
- Ratcliffe BC, Jameson ML, Smith ABT (2002) Scarabaeidae. In: Arnett American Beetles, Polyphaga: Scarabaeoidea through Curculionoidea, Ohio, USA.
- Leitão-Filho HF, Morellato LPC (1997) Semi deciduous forests of Southeastern Brasil ‒ Serra do Japi. In: Centres for plant diversity: a guide and strategy for their conservation. IUCN, WWF, Washington, USA.
- Oliveira-Filho AT, Ratter JA (2000) Padrões florísticos das matas ciliares da região do Cerrado e a evolução das paisagens do Brasil Central durante o Quaternário Tardio. In: Matas ciliares: conservação e recuperação. EDUSP, São Paulo.
- Endres AA, Creão-Duarte AJ, Hernández MIM (2007) Diversidade de Scarabaeidae s. str. (Coleoptera) da Reserva Biológica Guaribas, Mamanguape, Paraíba, Brasil: uma comparação entre Mata Atlântica e Tabuleiro Nordestino. Rev Bras Entomol 51: 67-71. Link: https://goo.gl/87XNm3
- Hernández MIM, Barreto PSCS, Valderêz HC, Creão-Duarte AJ (2014) Response of a dung beetle assemblage along a reforestation gradient in Restinga forest. J Insect Conserv 18: 539-546. Link: https://goo.gl/ziXq0T
- Medina AM, Lopes PP (2014) Seasonality in the dung beetle community in a Brazilian tropical dry forest: Do small changes make a difference? J Insect Sci 14: 1–11. Link: https://goo.gl/8yH3Pg
- McGeoch MA, Rensburg BJV, Botes A (2002) The verification and application of bioindicators: a case study of dung beetles in a savanna ecosystem. J Appl Ecol 39: 661-672. Link: https://goo.gl/8lDjdA
- Spector S, Ayzama S (2003) Rapid turnover and edge effects in Dung Beetle assemblages (Scarabaeidae) at a Bolivian Neotropical Forest–Savanna Ecotone. Biotropica 35: 394–404. Link: https://goo.gl/5dP7fc
- Gardner TA, Hernández MIM, Barlow J, Peres CA (2008) "Understanding the biodiversity consequences of habitat change: the value of secondary and plantation forests for neotropical dung beetles." J Appl Ecol 45:883-893. Link: https://goo.gl/VuG6Oj
- Sambrook J, Russel DW (2001) Molecular Cloning. 3rd edition. Cold Spring Harbor Laboratory Press, New York, USA. Link: https://goo.gl/GEc17T
- Rozen S, Skaletsky, H, (2000) Primer3 on the WWW for general users and for biologist programmers. In: Methods in Molecular Biology 132: 365-386. Link: https://goo.gl/gTgqzt
- Dieringer D, Schlötterer C (2003) Microsatellite Analyser (MSA): a platform independent analysis tool for large microsatellite data sets. Mol Ecol Notes 3: 167-169. Link: https://goo.gl/AQx6lS
- Roldán-Ruiz I, Dendauw J, Van BE, Depicker A, De Loose M (2000) AFLP markers reveals high polymorphic rates in ryegrasses (Lolium spp.). Mol Breed 6: 125–134. Link: https://goo.gl/F1diIy
- Excoffier L, Lisher HEL (2010) Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol Ecol Resour 10: 564–567. Link: https://goo.gl/Mwy7zY
- Van Oosterhout C, Hutchinson WF, Wills DPM, and Shipley P (2004) MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data. Mol Ecol Notes 4: 535-538.Link: https://goo.gl/GY1ii9
- Robertson HM (1993) The mariner transposable element is widespread in insects. Nature 362: 241-245. Link: https://goo.gl/G5Vlig






Table 1:
Characterization of four polymorphic microsatellite markers developed for C. (P) staigi (Scarabaeidae). Locus name, GenBank accession numbers, primer sequence (F: forward, R: reverse), anneling temperature (Ta), repeat motif, fragment size range in base pairs (bp), number of alleles per locus (A), observed heterozygosity (HO), expected heterozygosity (HE), and polymorphic information content (PIC).
Locus
GenBank
Ta (°C)
Repeat motif
Size range (bp)
A
HO
HE
PIC
Cstms01
KY794604
F: GGACTGTTTTGAGCGTTACCAT
R: GGCCCCTCCAGTATAGAAAATC
54.5
(TTGTG)2
161 - 196
5
0.533
0.572
0.312
Cstms02
KY794605
F: TTCTTATTGTAGCTTCGTTCATGC
R: GACTAACACATCTCACGGGGAA
52
(TGTCGA)2
397 - 361
5
0.499
0.583
0.339
Cstms04
KY794606
F: GCTGGTTTCATCAACTGCATTA
R: ACATACCACCCAGTCCTTTGTC
55.6
(TTATT)2
242-267
5
0.566
0.580
0.338
Cstms09
KY794607
F: AATCGCTTTAATCTTGTCTGGG
R: CTCAAAATCCCTAGAAGAACACG
55.6
(ATTT)3(ATGAAT)2
178 - 198
4
0.533
0.479
0.368